
![]()
Biopharmaceutical Sciences, Biomed Biopharm Res., 2021; 18(1):123-138
doi: 10.19277/bbr.18.1.256; download pdf version [+] here
Preliminary safety evaluation of n-butanol from the collagen extraction process and of collagen extract from Oreochromis niloticus (tilapia) skin oriented for dermocosmetics
1Universidad Nacional de Trujillo. Juan Pablo II Av. s/n, Trujillo, Perú; 2Department of Pharmacy, Faculty of Pharmaceutical Sciences, University of São Paulo, São Paulo, Brazil; 3Universidad de Los Lagos. Fuschlocher Av. 1305, Osorno, Chile
*corresponding authors:
Abstract
Obtaining new cosmetic ingredients, mainly from sustainable sources, as novel excipients or even active compounds is noteworthy for the cosmetic industry to enhance new innovative dermocosmetics. Thus, it is essential to establish the safety of these new ingredients to avoid adverse events, mainly those associated with clastogenic effects from the chemical compounds used for collagen extraction. In this study, we evaluated solutions of chemical compounds used in the collagen extraction process from tilapia skin (Oreochromis niloticus). The cytotoxic and genotoxic effects of the solutions used in the collagen extraction process were 10.0, 1.0, 0.5, and 0.1% n-butanol. Solutions were evaluated by the Allium test and the comet assay in peripheral white blood cells. The residual water from the final skin wash in the pre-treatment phase and the 0.5% lyophilized collagen extract were also investigated. The absence of cytotoxic and genotoxic activity was demonstrated in the collagen extract, despite the fact that n-butanol showed DNA damage, both in the root cells of Allium cepa and in the white blood cells of human peripheral blood. Therefore, we note the necessity to carry out genotoxicity tests to guarantee the absence of contaminants in the collagen extract for cosmetic purposes.
Keywords: Allium test; collagen extraction; Comet assay; cytotoxicity; genotoxicity; Oreochromis niloticus
Received: 26/02/2021; Accepted: 21/05/2021
Introduction
Sustainable use of residual materials from aquatic organisms has been widely studied during the last decade, with interesting results (1,2). Since the origin and the extraction method exert a strong influence on the purity and yield of collagen, there are many reviews available regarding marine resources that discuss not only the factors that influence the collagen extraction but also the physical and biological properties, such as its thermal stability, amino acid composition, and safety as oriented for dermocosmetics (1-4).
Moreover, numerous bioactive peptides of collagen have been extracted from fish via chemical or enzymatic hydrolysis as a safer option in the development of cosmeceutical products. Among the principal activities, we have examined the antimicrobial, antioxidant, antiphotoaging, photoprotective activities, and their potential application in the cosmetic area (2).
Indeed, there is a rising utilization of collagen from fish residuals for cosmetic applications since fish collagen has no risk in disease transmission to humans. For instance, some studies focused on wound healing applications with successful results as well as scar diminution (1,5–7). Fish collagen is principally derived from the skin of tilapia (Oreochromis niloticus) is highly appreciated for its biomedical and cosmetic uses due to particular physicochemical properties (8,9). The results of the biological evaluations of tilapia collagen demonstrated the absence of cellular toxicity and chromosomal aberrations, negative sensitivity tests, and acute systemic toxicity (10). Biocompatibility and regenerative activity in the treatment of deep second-degree burns have also been observed (11). The biological safety of collagen will depend on its purity; therefore, it is necessary to assess pretreatment procedures and methods of extraction.
The methods for extracting collagen from by-products of different animal origins are well established (10,12). To release non-collagen proteins, fats, and pigments, sodium hydroxide and n-butanol in proportions of 1:20 weight/volume (w/v) are commonly used in the first stage of treatment (13,14). After treatments at variable times with these chemical compounds, successive rinses are carried out to remove chemical residues infused within the extracted collagen or in the rinsing liquids. Some of these molecules, such as hydrogen peroxide (15) and n-butanol (16), represent a source for the environmental pollution and potential genotoxicity.
Biosafety of raw materials in cosmetic industries is mandatory. It is also important to ensure that the concentrations of chemical substances used during extraction processes for cosmetic (ingredient) use must be eliminated or reduced (17). For collagen extraction, caution in the elimination of any remaining n-butanol in the by-products of the collagen raw material (14) is required.
Even when an absence of butanol mutations is reported using the Ames assay (16), additional tests are needed to confirm the results and to specify the utilized concentrations. Current research shows that a single assay is not sufficient to exclude a genotoxic substance, even when the result has been negative (17–20). In this research, we evaluated the safety of chemical solutions of n-butanol (used in the extraction process), residual water from washes and collagen extract from O. niloticus skin itself with cosmetic purposes, using cytotoxic and genotoxic assays by Allium and comet tests.
Materials and Methods
Biological material
1) Allium cepa bulbs, medium in size, with germinal disk between 1 and 2 cm.
2) Peripheral blood lymphocytes obtained from two healthy male donors, aged 25 and 38 years. The donors were non-smokers and did not consume drugs. (N°: 001-2020/CEI-UNT).
3) Tilapia (Oreochromis niloticus), 12 months old (30 fish, 450 ± 35.0 g).
Chemical products
n-Butanol (1-butanol CAS 71-36-3), NaOH, acetic acid, hydrogen peroxide, NaCl, disodium EDTA, Tris HCl, Triton X-100 and DMSO from Sigma-Aldrich were used. Propidium iodide and SYBR Gold from Invitrogen, USA were also used.
Obtaining collagen from tilapia (O. niloticus)
Collagen was obtained by extraction in acetic acid (ASC), according to a recent report (9). After successive washes with an abundance of distilled water, the tilapia skin was pre-treated with 0.1M NaOH (1:20 w/v) for 24 h, followed by treatment with n-butanol at 10% (1:20 w/v) . Ten washes with an abundance of distilled water were performed, followed by collagen extraction in 0.5M acetic acid for 48 h. The entire procedure was conducted at 4.0 °C, with recovered collagen vacuum filtered with a filter paper (Whatman N° 4), and then lyophilized.
Allium test
The onion bulbs were immersed in water, confirming that the germinal disk was fully immersed. Testing was initiated when (all of) the roots reached between 1 and 2 cm in length. Successive tests of n-butanol at concentrations of 10.0; 1.0; 0.5; 0.1; 0.05 and 0.025% were performed to determine both viability and activity of the cells. The growing roots were then exposed to either n-butanol, residual water from the final (10th) rinse of the pretreatment of the collagen extraction process, 0.5% collagen in distilled water, and purified water, as a control. After 6 h of treatment, the apical meristems were cut from the roots, fixed in an ethanol:acetic acid solution (3:1 v/v) and held at -4.0 °C for a minimum of 24 h. Samples were stained with 1.0% acetic orcein 1N: HCl (9: 1 v/v) for 30 minutes (21). The samples were crushed and observed with an optical microscope at 40 and 100X. Three replicates were performed.
Comet assay
Ten mL of peripheral blood were collected from the volunteers. White blood cells were immediately isolated and exposed to n-butanol (0.01 and 0.001%), 0.5% tilapia collagen solution, PBS (negative control) or 12% hydrogen peroxide (positive controls). After 2 h of treatment, the white blood cells viability was determined by staining with 1.0 mM propidium iodide (Invitrogen, USA) and controlled with an Olympus BX51 40X fluorescence microscope using the 536 and 617 nm filters. The samples were placed on slides previously prepared with a layer of agarose and cell lysis was performed with the buffer containing NaCl 2.5M, Na2EDTA 100mM, Tris HCl 10mM, Triton X-100 1%, and DMSO 10%. The samples were then incubated at 4.0 °C for 4 h, followed by electrophoresis at 25V (Power Pack P25 Timer, Biometra GmbH, Germany) and 300mA for 30 minutes in buffer pH 13 (1.0mM Na2EDTA and 300mM NaOH). The samples were neutralized in Tris HCl solution pH 7.5 for 3 minutes and fixed in 96° alcohol for 3 minutes (22). Samples were stained with SYBR Gold and observed under a fluorescence microscope with 40X magnification (23).
Data collection
The frequency of the cell cycle phases was recorded and typified in the different treatments and in the treatments that registered genetic changes in the Allium test. The DNA damage was quantified using the Score Assay program and the percentage of DNA in the tail (% Tail DNA) was considered a genotoxicity parameter in the comet assay (24).
Statistical Analysis
The averages of each variable were compared by analysis of variance with the Origin Pro 2020 program (OriginLab Corporation, USA) and regression analysis was applied to reveal the dose-response relationship of chemical compounds, with a reliability level of α = 0.05.
Results and Discussion
Collagen extraction methods are well described in the literature. In general, three steps are commonly used to extract low molecular weight collagen hydrolysates from natural resources: initial treatment of the raw material, gelatin (collagen) extraction, and collagen hydrolysis. In the pretreatment (initial treatment) phase, sodium hydroxide is used to eliminate non-collagen proteins and butyl alcohol used to reduce lipids. The last step could be divided into enzymatic, chemical, and subcritical water hydrolysis (3).
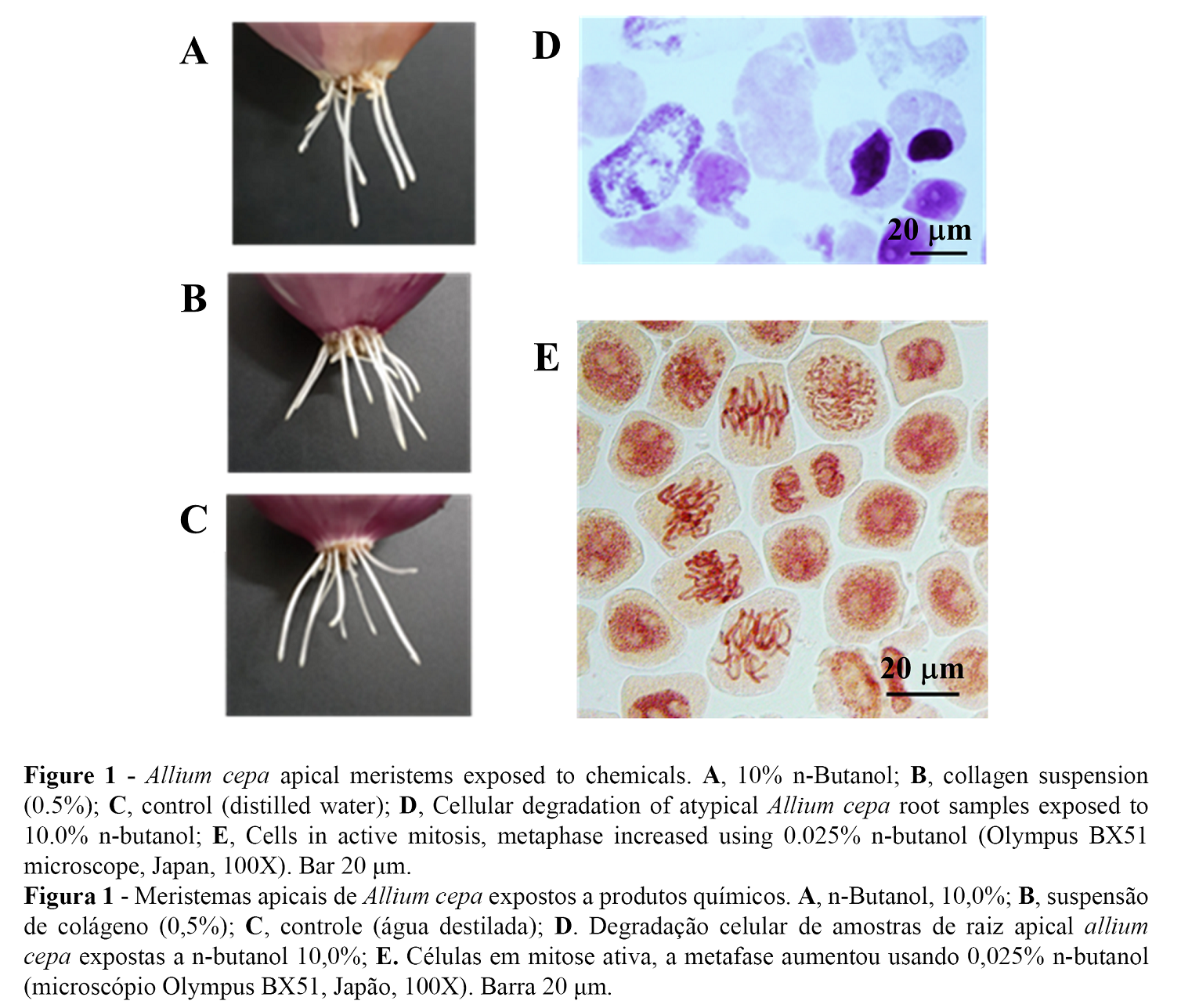
Dynamics of cell proliferation in the apical meristem of Allium cepa, achieved between 1 to 3 days of growth induction is a sensitive biological scenario to detect substances that can affect the genetic material (25). Thus, Allium test is comparable to assays on animals and cell line cultures (26). As shown in Figure 1A, we observed transparent aspect and withered roots, unlike the roots shown in Figures 1B and 1C, which corresponded to the treatments with the collagen suspension (0.5%) and control growth in distilled water, respectively. Degradation of the nucleus and cytoplasm were observed in the samples exposed to 10.0% n-butanol for 6 h (Figure 1D), thus this concentration can be considered lethal. Moreover, high inhibition of cell activity was observed at 1.0% n-butanol, mitotic index of 2.99% with absence of normal cells in the metaphase; and the mitotic index of 2.75% (not significant) was found at 0.5% n-butanol with the presence of mitotic phases. In addition, there were four metaphases, two prophases and one telophase in an unusual single visual field at the concentration of 0.025% n-butanol (Figure 1E).
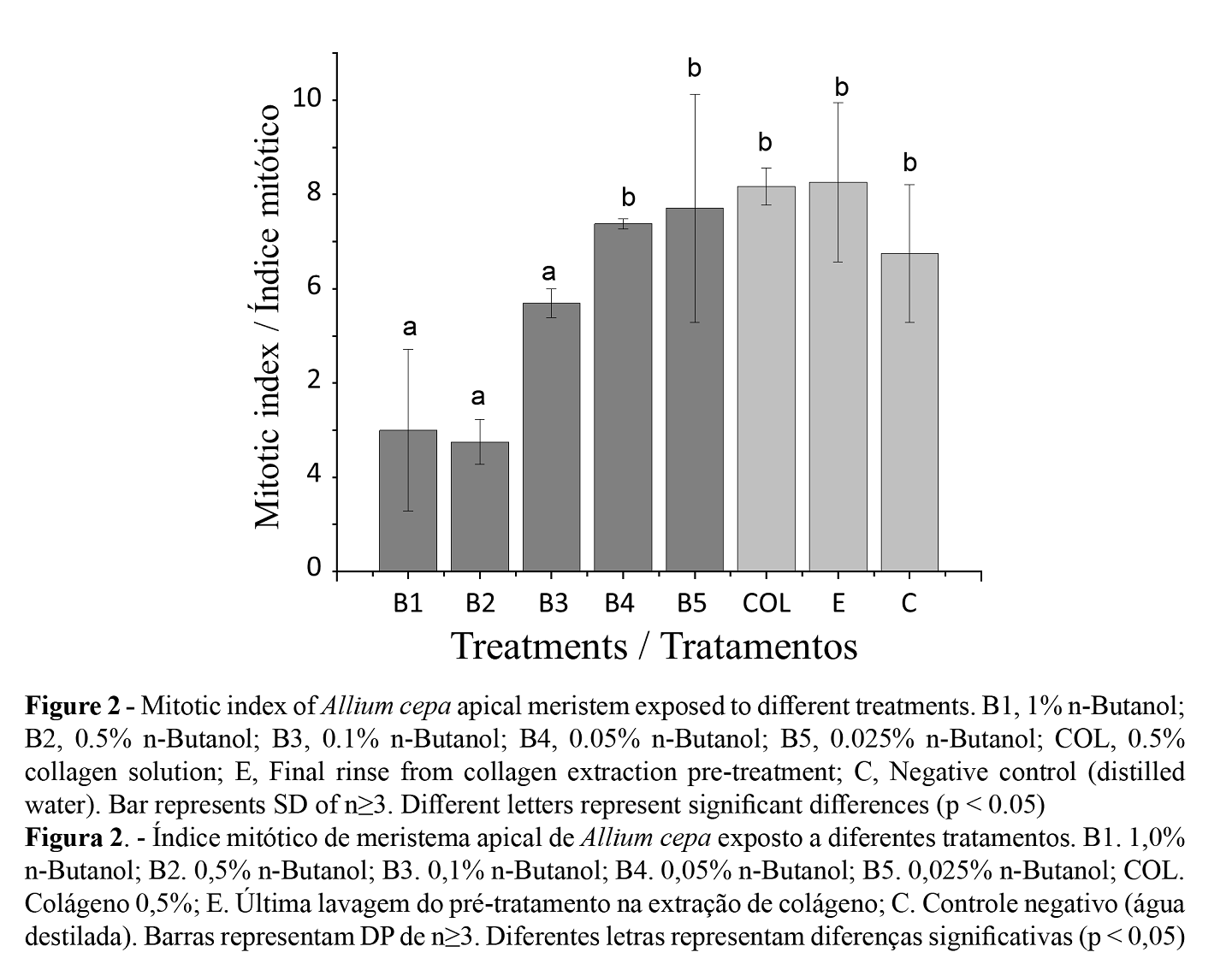
The percentage of cell divisions in root meristems of A. cepa was determined by the mitotic index. Lethality and no cell activity were observed with 10.0% n-butanol, while for 1.0, 0.5 and 0.1% n-butanol, a significant decrease in cell activity was observed compared to those treated with lower concentrations (0.05 and 0.025%). These last indices were compared with those of the negative control, collagen solution at 0.5% and the residual water of the final (tenth) rinse of the pretreatment process (Figure 2).
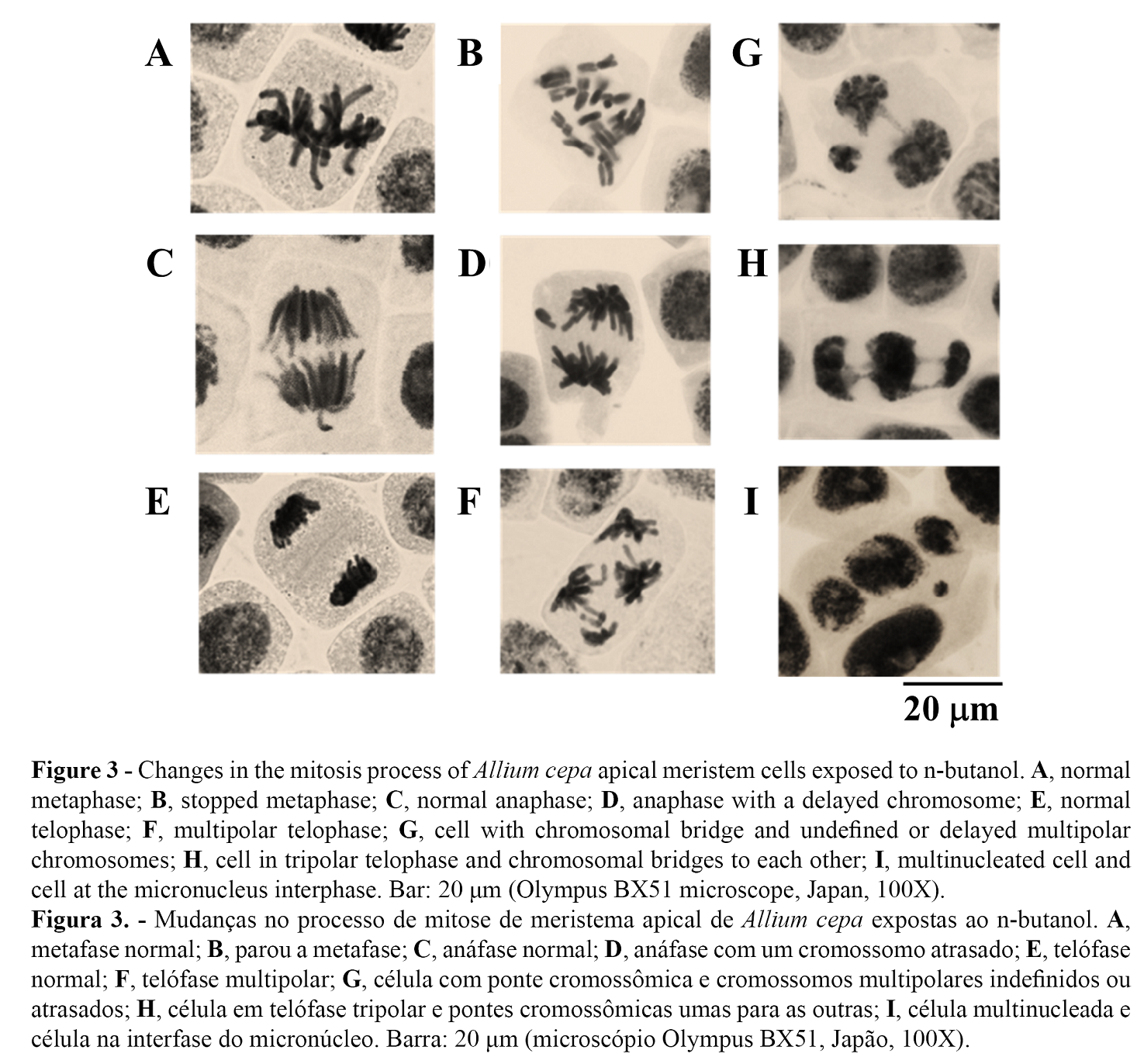
We observed the following cellular level changes produced by n-butanol in A. cepa at the metaphase level: partial cytostatic effect at concentrations of 0.5 and 1.0%. Figure 3A shows normal metaphase and Figure 3B shows stopped metaphase (c-metaphase), in which the chromosomes were dispersed and highly condensed. This irregular variation was an indicative of the absence or failure of the achromatic spindle formation, as well as signs of chromosomal compaction due to metaphasic stagnation (27). Variants of the normal phases of anaphase and bipolar telophase (Figures 3C and 3E, respectively) in comparison with the presence of lag chromosomes in anaphase (Figure 3D) and multipolar structure in telophase (Figure 3F), are evidence to contribute to the dysfunction of the achromatic spindle and defective centrosome division, as illustrated in Figure 3F, with four polar centers rather than two. The assembly of the mitotic spindle organized in a bipolar way, with dynamic balance of kinesin-5 and dynein, ensures equitable distribution of chromosomes during segregation in mitosis (28).
Furthermore, the dysfunctional organization in supernumerary centrosomes leads to flaws in cytokinesis and numerical variations of chromosomes in the resulting cells (29). Figure 3G-I shows another alteration produced by n-butanol was the breakdown of chromosomes. Figure 3G illustrates a telophase cell with a chromosome bridge; moreover, not clearly defined whether they were delayed or tripolar chromosomes. In the absence of cytokinesis, a tripolar cell with chromosome bridges between chromosome groups would increase to a cell with three nuclei, as in Figure 3H,I. Likewise, the presence of micronuclei (Figure 3I) indicated chromosomal breaks produced in phases prior to mitosis.
The alterations recorded after 6 h of exposure were registered at the final stage of replication process (S phase) and G2 phase (premitotic), as well as the presence of multipolar-centers. They can occur as a consequence of the cellular and chromosomal cytoskeleton instability due to the effect of n-butanol, which would have produced fragmentation of the pre-existing centrosome(s) instead of a centrosome amplification. The formation of multipolar spindle regardless of the amplification of the centrosome in HeLa cells after irradiation in G2 phase has been previously observed (30). Moreover, it is known that the centrosome duplication has occurred only once in each cell cycle and began in the G1 (pre-replication)/S phase and completed during G2 phase (31). However, future investigations with a centrosomal labeling should be carried out to determine if it is a fragmentation or amplification.
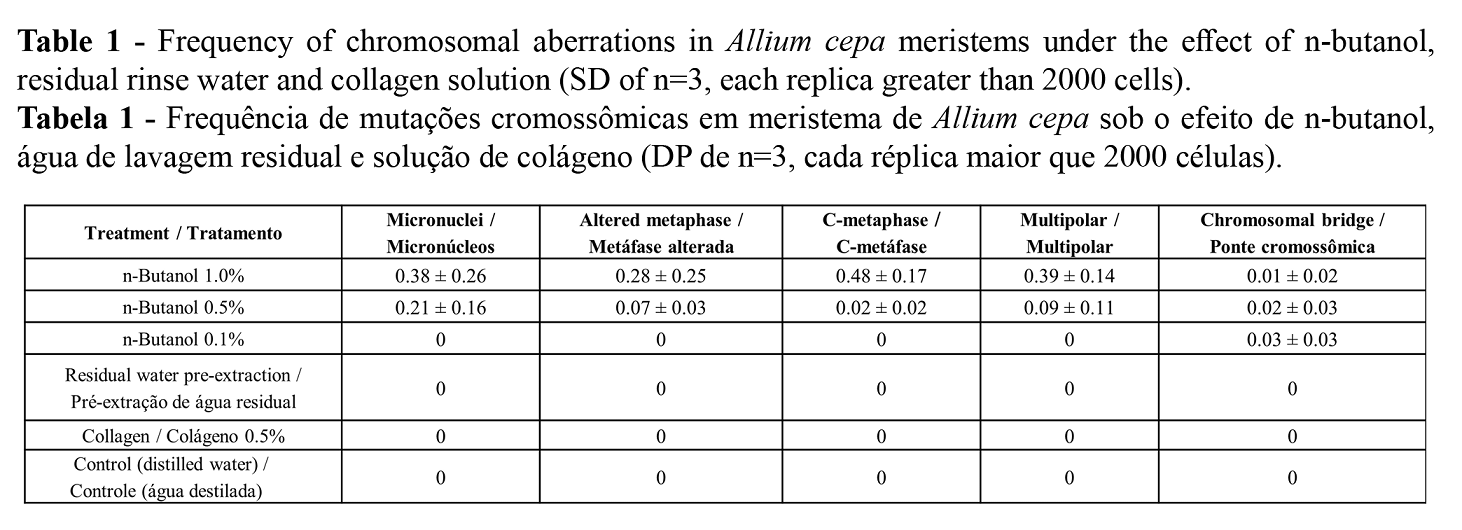
The quantified changes due to the effect of n-butanol are summarized in Table 1. Multipolar cells were most frequently observed, followed by metaphases with dispersed and condensed chromosomes. Chromosomal changes were not observed in the other treatments, thus verifying the genotoxicity of n-butanol.
The comet assay is one of the methods used to measure DNA damage, using whole blood (22) or white blood cells isolated directly from peripheral blood (5). Among the parameters of genotoxicity, the percentage of DNA in the comet test is used as the most informative parameter due to its relationship with the magnitude of the DNA damage. The greater the DNA breakage, the greater the tail length due to migration, therefore, the visualization will be immediate and the intensity of the tail more evident (32). DNA damage was quantified using the DNA percentage parameter on the tail.
In our investigation, the toxicity of n-butanol was determined by the percentage of living cells with the differential staining given by propidium iodide in white blood cells isolated from human peripheral blood. Only dead cells were colored, as seen in Figure 4. Concentrations above 0.1% n-butanol were lethal in the white blood cells. The percentage of viability of white blood cells exposed to 0.001% n-butanol was 96.87%. Cellular viability greater than 75% has been reported to be necessary for the validity of genotoxicity tests (33).
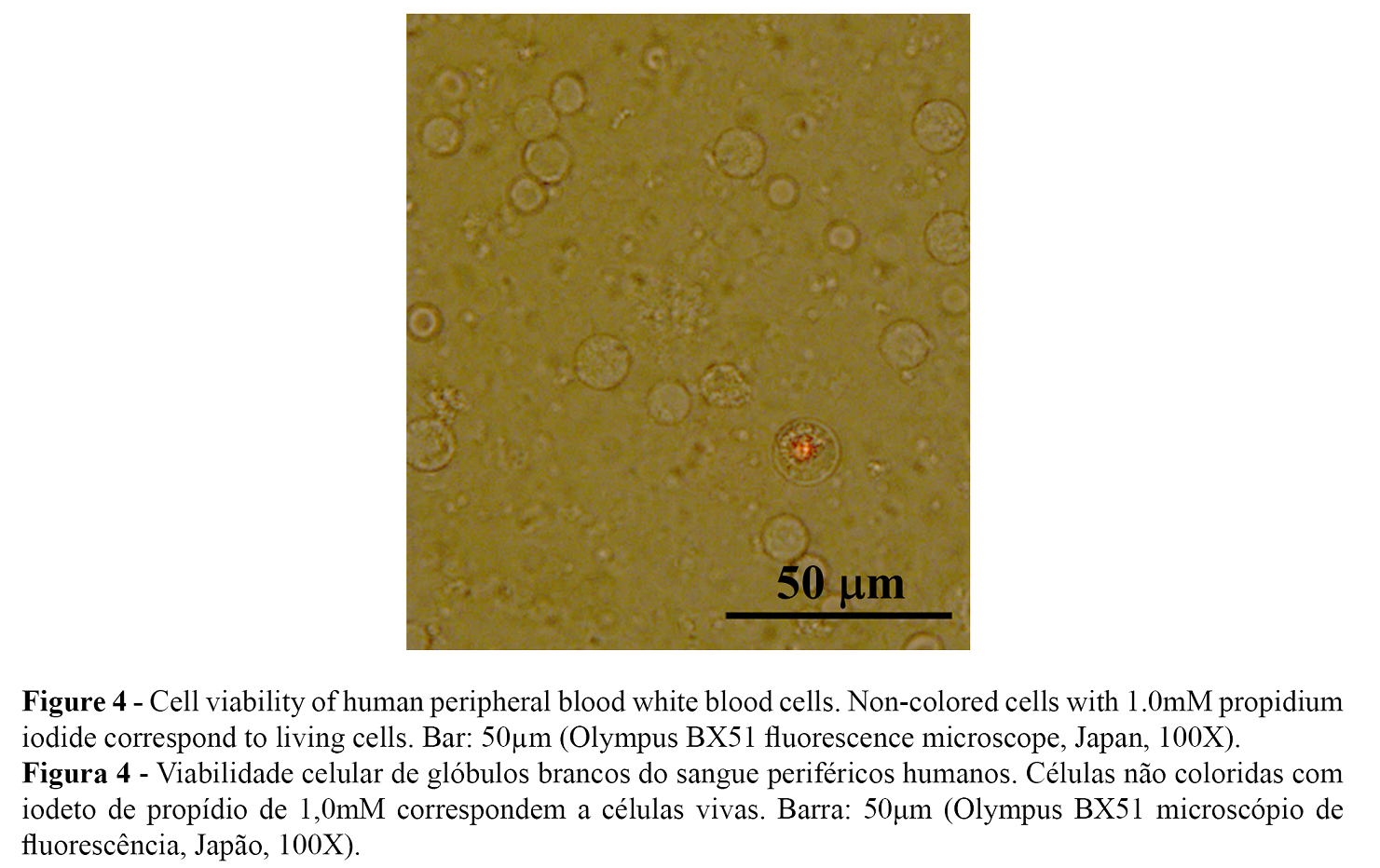
DNA damage of white blood cells due to the exposure to n-butanol was demonstrated by the degree of DNA migration in alkaline unicellular electrophoresis (pH> 13). The heterogeneity of the response of white blood cells to a clastogenic compound was observed at the same sub-lethal dose and exposure time recorded in all investigations (34). Figure 5 shows the variable magnitude of DNA damage in white blood cells, from zero level (no damage) in A, mild damage levels in B and C, medium damage in D and high damage in E and F. These types of damage were recorded in all treatments including the positive and negative controls, using the % Tail DNA to determine the clastogenic activity.
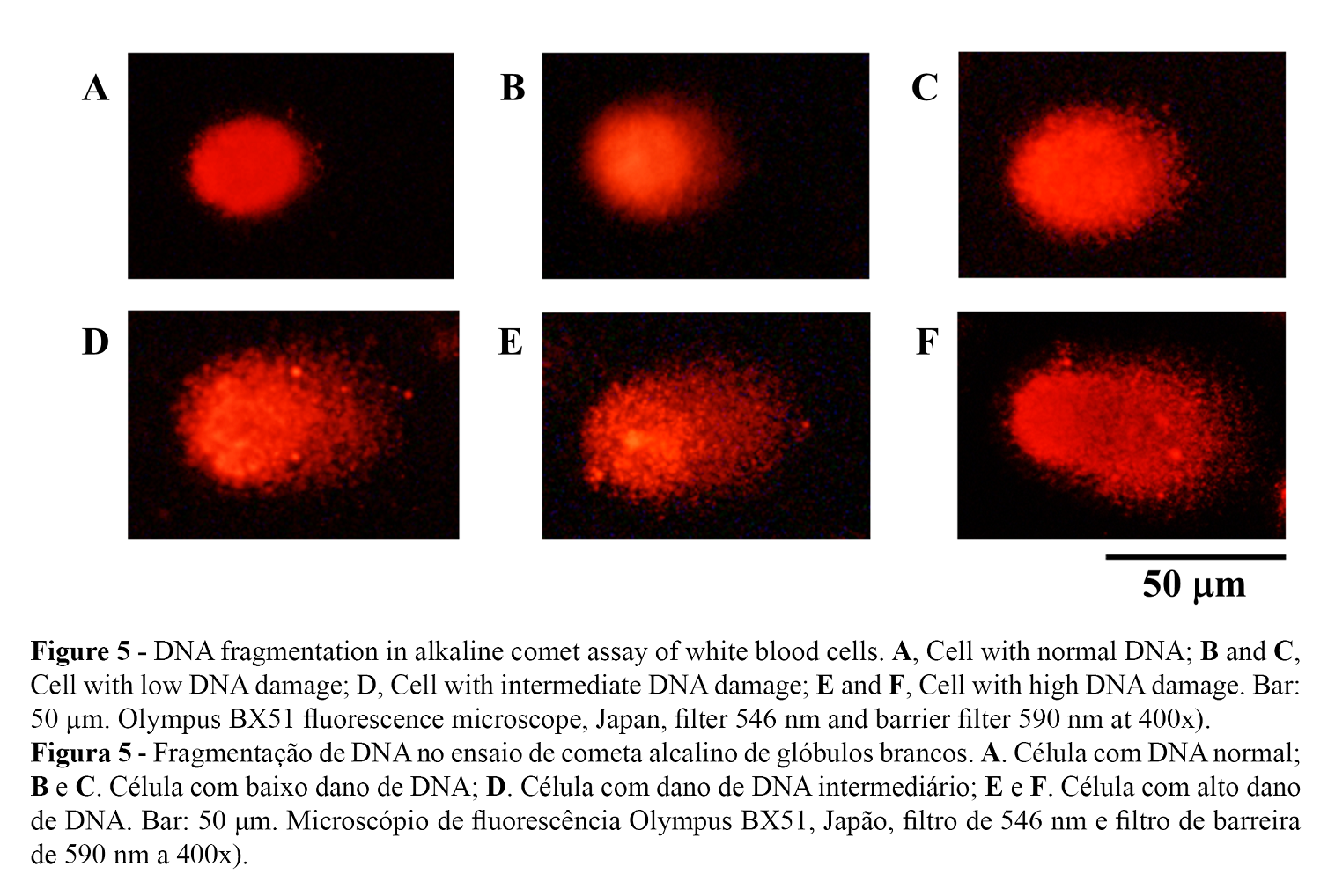
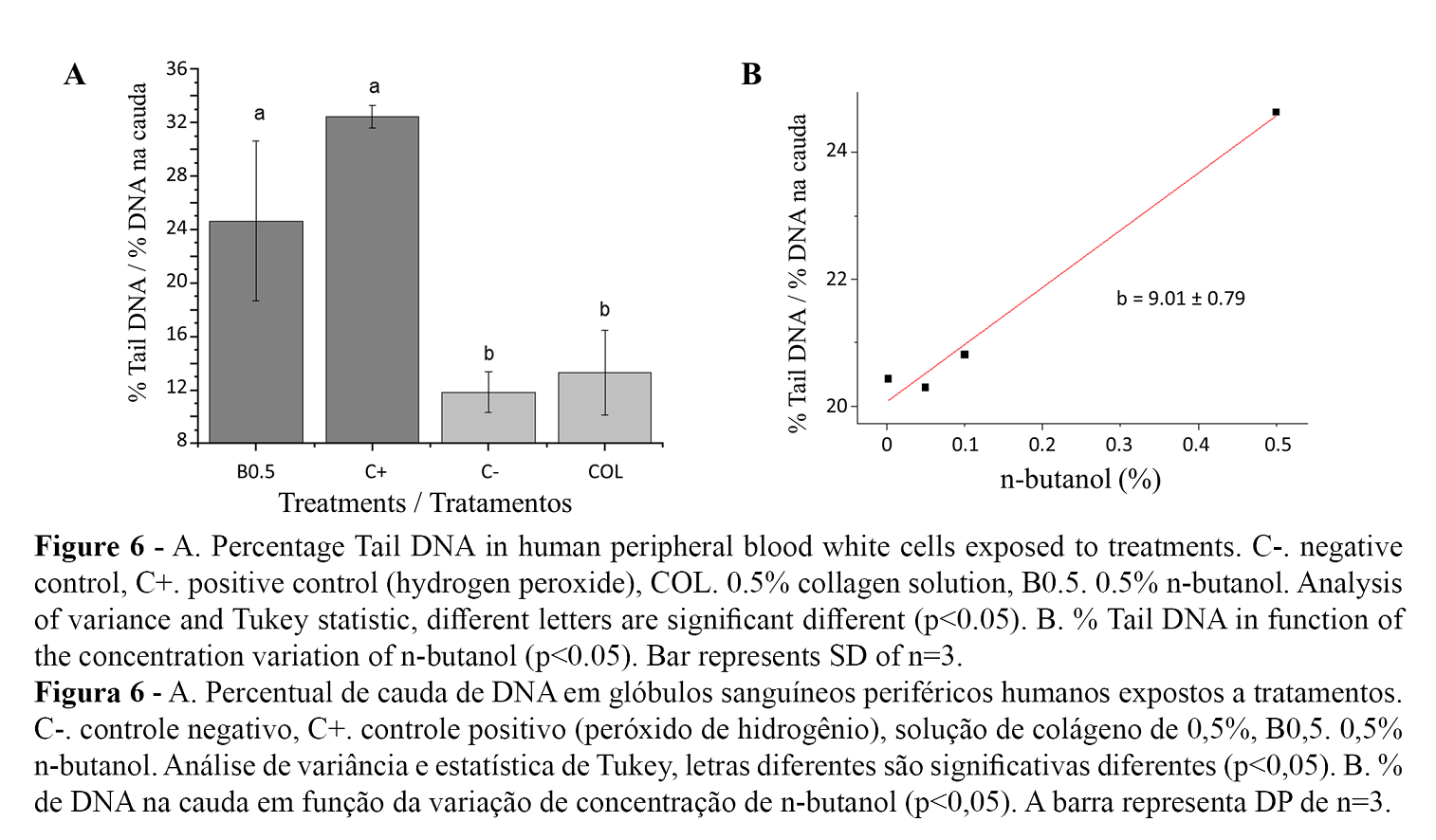
Figure 6A. shows the comparison of averages of the DNA percentage in the comet tail for the treatment with n-butanol (0.5%), without differences with the positive control and significantly different to the negative control, as well as the collagen extract (0.5%). The percentage of viability evaluated after the n-butanol treatment remained above 95% with an average between treatments of 97.20 ± 0.12%. Concentrations lower than 0.1% n-butanol showed a decrease in the clastogenic effect without significant variations with concentration (Figure 6B). The high variability in the damage degree of the white blood cells exposed to n-butanol within each treatment reflects the variation in the sensitivity of cellular response.
In the cytological tests of A. cepa with the rinsing liquid and with 0.5% collagen, the mitotic cycle was normal with no evidence of chromosomal damage. If the rinses were insufficient, chemical residues could remain in the collagen. Fortunately, due to the low-fat content in tilapia skin (3.85%) (35), compared to other species of fish, such as Sardinella maderensis, (26.0%) and S. aurita, (24.0%) (36), n-butanol could be replaced by ethyl alcohol to remove fat (37). Another alternative treatment to eliminate fat from the raw material is fermentation with Bacillus velezensis, which has a high capacity to metabolize fat and proteins (38).
It has been shown that 1-butanol or n-butanol is absorbed through the skin and respiratory tract and produces pathological changes in the liver and kidneys in rats, but not a mutagenic or carcinogenic effect (39). Likewise, investigations in pregnant rats exposed to 1.0% n-butanol administered through water reported toxicity and a decrease in maternal and fetal body weight (40). For the Ames test, non-mutagenicity was reported for n-butanol (16). In contrast, zebrafish embryos exposed to n-butanol exhibited severe oxidative damage with consequent damages in liver and brain tissues, according to a recent study (41). However, our study revealed the possibility of genetic changes if n-butanol is continuously used for long periods of time.
In summary, we revealed the DNA fragmentation of interphase, specialized and viable cells exposed in vitro to n-butanol with the comet assay. Moreover, alterations in addition to DNA breaks were detected with the Allium test. However, further study is required to provide information about the mechanisms of action of the n-butanol DNA damage.
Conclusions
In the safety assessment of collagen extract obtained from the tilapia (Oreochromis niloticus) skin for cosmetic use, we concluded that n-butanol was a genotoxic compound used, with significant changes in the mitotic apparatus and chromosomes of Allium cepa as well as a high percentage of DNA in the tail of the comets. In addition, we revealed clear evidence to consider partial (n-butanol) replacement or the verification of its complete elimination with successive (water) washes to minimize its presence in the raw material for further preparations containing natural products, such as this collagen derived from sustainable sources. We note the importance of providing multiple proofs of the genotoxic activity of the chemicals used in the collagen extraction process to ensure the absence of potentially harmful contaminants in the materials to be used within cosmetic formulations.
Authors Contributions Statement
Conceptualization, Z.A.P., C.A.-J.; methodology, L.S.-T., N.R.-P.; formal analysis, A.R.B., D.S., C.A.-J., R.F.; investigation, Z.A.P., R.Q.-L., A.R.B., A.L.M.-J., C.A.-J., D.S.; writing - original draft preparation, Z.A.P., A.L.M.-J., F.D.S., A.R.B.; writing—review and editing, A.R.B., R.M.M., A.L.M.-J.; supervision, Z.A.P., C.A.-J.; project administration, Z.A.P., C.A.-J.; funding acquisition, Z.A.P., R.Q.-L., A.R.B. All authors have read and agreed to the published version of the manuscript.
Funding
Authors are very thankful to CONCYTEC-FONDECYT, process E041-01 (Contract number 123-2018-FONDECYT-BM-IADT/AV), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) Process 305250/2019-1, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brasil (CAPES) Finance Code 001 and Programa Nacional de Pós-Doutorado (PNPD).
Acknowledgments
The authors are very thankful to Mónica Arqueros, biologist, for her help with the Comet Test.
Conflicts of Interest
The authors declare no conflict of interest.
Ethical Statement
This research was developed under the Ethical Process N°: 001-2020/CEI-UNT (Universidad de Trujillo).
References
- Salvatore, L., Gallo, N., Natali, M. L., Campa, L., Lunetti, P., Madaghiele, M., Blasi, F. S., Corallo, A., Capobianco, L., & Sannino, A. (2020).Marine collagen and its derivatives: Versatile and sustainable bio-resources for healthcare. Materials science & engineering. C, Materials for biological applications, 113,110963. https://doi.org/10.1016/j.msec.2020.110963
- Venkatesan, J., Anil, S., Kim, S. K., & Shim, M. S. (2017). Marine Fish Proteins and Peptides for Cosmeceuticals: A Review. Marine drugs, 15(5), 143. https://doi.org/10.3390/md15050143
- Hong, H., Fan, H., Chalamaiah, M., & Wu, J. (2019). Preparation of low-molecular-weight, collagen hydrolysates (peptides): Current progress, challenges, and future perspectives. Food chemistry, 301,125222. https://doi.org/10.1016/j.foodchem.2019.125222
- Halim, N.R., Yusof, H., & Sarbon, N.M. (2016). Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends in Food Science and Technology, 51, 24-33. doi:10.1016/j.tifs.2016.02.007
- Ge, B., Wang, H., Li, J., Liu, H., Yin, Y., Zhang, N., & Qin, S. (2020). Comprehensive Assessment of Nile Tilapia Skin (Oreochromis niloticus) Collagen Hydrogels for Wound Dressings. Marine drugs, 18(4), 178. https://doi.org/10.3390/md18040178
- Mei, F., Liu, J., Wu, J., Duan, Z., Chen, M., Meng, K., Chen, S., Shen, X., Xia, G., & Zhao, M. (2020). Collagen Peptides Isolated from Salmo salarand Tilapia niloticaSkin Accelerate Wound Healing by Altering Cutaneous Microbiome Colonization via Upregulated NOD2 and BD14. Journal of agricultural and food chemistry, 68(6), 1621–1633. https://doi.org/10.1021/acs.jafc.9b08002
- Aguirre-Cruz, G., León-López, A., Cruz-Gómez, V., Jiménez-Alvarado, R., & Aguirre-Álvarez, G. (2020).Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants (Basel, Switzerland), 9(2), 181. https://doi.org/10.3390/antiox9020181
- Liao, W., Guanghua, X., Li, Y., Shen, X. R., & Li, C. (2018). Comparison of characteristics and fibril-forming ability of skin collagen from barramundi (Lates calcarifer) and tilapia (Oreochromis niloticus). International journal of biological macromolecules, 107(Pt A), 549–559. https://doi.org/10.1016/j.ijbiomac.2017.09.022.
- Li, J., Wang, M., Qiao, Y., Tian, Y., Liu, J., Qin, S., & Wu, W. (2018). Extraction and characterization of type I collagen from skin of tilapia (Oreochromis niloticus) and its potential application in biomedical scaffold material for tissue engineering. Process Biochemistry, 74, 156-163. https://doi.org/10.1016/j.procbio.2018.07.009.
- Yamamoto, K., Igawa, K., Sugimoto, K., Yoshizawa, Y., Yanagiguchi, K., Ikeda, T., Yamada, S., & Hayashi, Y. (2014). Biological safety of fish (tilapia) collagen. BioMed research international, 2014, 630757. https://doi.org/10.1155/2014/630757
- Pati, F., Datta, P., Adhikari, B., Dhara, S., Ghosh, K., & Das Mohapatra, P. K. (2012).Collagen scaffolds derived from fresh water fish origin and their biocompatibility. Journal of biomedical materials research. Part A, 100(4), 1068–1079. https://doi.org/10.1002/jbm.a.33280
- Hong, H., Fan, H., Roy, B. C., & Wu, J. (2021). Amylase enhances production of low molecular weight collagen peptides from the skin of spent hen, bovine, porcine, and tilapia. Food chemistry, 352,129355. https://doi.org/10.1016/j.foodchem.2021.129355
- Sun, L., Hou, H., Li, B., & Zhang, Y. (2017). Characterization of acid- and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). International journal of biological macromolecules, 99, 8–14. https://doi.org/10.1016/j.ijbiomac.2017.02.057.
- Liu, W., Zhang, Y., Cui, N., & Wang, T. (2019). Extraction and characterization of pepsin-solubilized collagen from snakehead (Channa argus) skin: Effects of hydrogen peroxide pretreatments and pepsin hydrolysis strategies. Process Biochemistry, 76,194-202 doi:10.1016/j.procbio.2018.10.017.
- Benhusein, G. M., Mutch, E., Aburawi, S., & Williams, F. M. (2010). Genotoxic effect induced by hydrogen peroxide in human hepatoma cells using comet assay. The Libyan journal of medicine, 5,10.3402/ljm.v5i0.4637. https://doi.org/10.3402/ljm.v5i0.4637.
- McCann, J., Choi, E., Yamasaki, E., & Ames, B. (1975). Detection of Carcinogens as Mutagens in the Salmonella/Microsome Test: Assay of 300 Chemicals. Proceedings of the National Academy of Sciences of the United States of America, 72(12), 5135-5139. http://www.jstor.org/stable/65363
- Demir, Y., Uckaya, M., & Demir, N. (2019).Evaluation of the efficacy in cosmetic products safety: Comparison with biochemical substrates. Regulatory toxicology and pharmacology : RTP, 104, 56–58. https://doi.org/10.1016/j.yrtph.2019.03.001
- Prantl, E. M., Kramer, M., Schmidt, C. K., Knauer, M., Gartiser, S., Shuliakevich, A., Milas, J., Glatt, H., Meinl, W., & Hollert, H. (2018). Comparison of in vitro test systems using bacterial and mammalian cells for genotoxicity assessment within the "health-related indication value (HRIV) concept. Environmental science and pollution research international, 25(5), 3996–4010. https://doi.org/10.1007/s11356-016-8166-z
- Fiskesjö G. (1985). The Allium test as a standard in environmental monitoring. Hereditas, 102(1), 99–112. https://doi.org/10.1111/j.1601-5223.1985.tb00471.x
- Collins, A., Koppen, G., Valdiglesias, V., Dusinska, M., Kruszewski, M., Møller, P., Rojas, E., Dhawan, A., Benzie, I., Coskun, E., Moretti, M., Speit, G., Bonassi, S., & ComNet project (2014). The comet assay as a tool for human biomonitoring studies: the ComNet project. Mutation research. Reviews in mutation research, 759, 27–39. https://doi.org/10.1016/j.mrrev.2013.10.001
- Giménez-Abián, M.I., Torre, C., & López-Sáez, J.F. (1987).Growth and cell proliferation in Allium roots at different oxygen tensions. Environmental and Experimental Botany, 27, 233-237..
- Azqueta, A., & Collins, A. R. (2013). The essential comet assay: a comprehensive guide to measuring DNA damage and repair.Archives of toxicology, 87(6), 949–968. https://doi.org/10.1007/s00204-013-1070-0.
- Amésquita, L., Cruz-Briceño, M. N., & Prieto, Z. (2018). Daño en el ADN de linfocitos humanos por efecto de cloroquina [Damage to human lymphocyte DNA from chloroquine effect]. Revista peruana de medicina experimental y salud publica, 35(3), 471–475. https://doi.org/10.17843/rpmesp.2018.353.3166
- Reisinger, K., Blatz, V., Brinkmann, J., Downs, T. R., Fischer, A., Henkler, F., Hoffmann, S., Krul, C., Liebsch, M., Luch, A., Pirow, R., Reus, A. A., Schulz, M., & Pfuhler, S. (2018). Validation of the 3D Skin Comet assay using full thickness skin models: Transferability and reproducibility. Mutation research. Genetic toxicology and environmental mutagenesis, 827,27–41. https://doi.org/10.1016/j.mrgentox.2018.01.003
- Calvo, A., Cruz, J.L., Gutiérrez, C., García-Herdugo, G., Giménez-martin, G., & López-Sáez, J.F. (1982).Analysis of cell cycle in root meristems.Journal of Theoretical Biology, 96, 295-308. https://doi.org/10.1016/0022-5193(82)90226-0.
- Bonciu, E., Firbas, P., Fontanetti, C.S., Wusheng, J., Karaismailoğlu, M.C., Liu, D., Menicucci, F., Pesnya, D.S., Popescu, A., Romanovsky, A., Schiff, S., Ślusarczyk, J., Souza, C.P., Srivastava, A., Șuțan, A., & Papini, A. (2018). An evaluation for the standardization of the Allium cepa test as cytotoxicity and genotoxicity assay. Caryologia, 71, 191 - 209. doi:10.1080/00087114.2018.1503496
- Thadani, R., Kamenz, J., Heeger, S., Muñoz, S., & Uhlmann, F. (2018).Cell-Cycle Regulation of Dynamic Chromosome Association of the Condensin Complex. Cell reports, 23(8), 2308–2317. https://doi.org/10.1016/j.celrep.2018.04.082.
- Batty, P., & Gerlich, D. W. (2019). Mitotic Chromosome Mechanics: How Cells Segregate Their Genome. Trends in cell biology, 29(9), 717–726. https://doi.org/10.1016/j.tcb.2019.05.007
- Leone, M., & Engel, F. B. (2019).Pseudo-bipolar spindle formation and cell division in postnatal binucleated cardiomyocytes. Journal of molecular and cellular cardiology, 134,69–73. https://doi.org/10.1016/j.yjmcc.2019.07.005
- Fletcher, L., & Muschel, R. J. (2006). The centrosome and the DNA damage induced checkpoint. Cancer letters, 243(1), 1–8. https://doi.org/10.1016/j.canlet.2006.01.006
- Tanaka K. (2014). Centrosome duplication: suspending a license by phosphorylating a template. Current biology : CB, 24(14), R651–R653. https://doi.org/10.1016/j.cub.2014.06.008.
- Collins, A. R., Oscoz, A. A., Brunborg, G., Gaivão, I., Giovannelli, L., Kruszewski, M., Smith, C. C., & Stetina, R. (2008). The comet assay: topical issues.Mutagenesis, 23(3), 143–151. https://doi.org/10.1093/mutage/gem051
- Henderson, L., Wolfreys, A., Fedyk, J., Bourner, C., & Windebank, S. (1998). The ability of the Comet assay to discriminate between genotoxins and cytotoxins. Mutagenesis, 13(1), 89–94. https://doi.org/10.1093/mutage/13.1.89
- Olive, P. L., & Durand, R. E. (2005).Heterogeneity in DNA damage using the comet assay. Cytometry. Part A : the journal of the International Society for Analytical Cytology, 66(1), 1–8. https://doi.org/10.1002/cyto.a.20154
- Alfaro, A.T., Fonseca, G., Balbinot, E., Machado, A.G., & Prentice, C. (2013).Physical and chemical properties of wami tilapia skin gelatin. Food Science and Technology International, 33,592-595.doi.org/10.1590/S0101-20612013005000069
- Njinkoué, J. M., Barnathan, G., Miralles, J., Gaydou, E. M., & Samb, A. (2002). Lipids and fatty acids in muscle, liver and skin of three edible fish from the Senegalese coast: Sardinella maderensis, Sardinella aurita and Cephalopholis taeniops. Comparative biochemistry and physiology. Part B, Biochemistry & molecular biology, 131(3), 395–402. https://doi.org/10.1016/s1096-4959(01)00506-1
- Menezes, M., Ribeiro, H. L., Abreu, F., Feitosa, J., & Filho, M. (2020).Optimization of the collagen extraction from Nile tilapia skin (Oreochromis niloticus) and its hydrogel with hyaluronic acid.Colloids and surfaces. B, Biointerfaces, 189, 110852. https://doi.org/10.1016/j.colsurfb.2020.110852
- Song, Z., Liu, H., Chen, L., Chen, L., Zhou, C., Hong, P., & Deng, C. (2021). Characterization and comparison of collagen extracted from the skin of the Nile tilapia by fermentation and chemical pretreatment. Food chemistry, 340,128139. https://doi.org/10.1016/j.foodchem.2020.128139
- International Programme on Chemical Safety, United Nations Environment Programme, International Labour Organization & World Health Organization. (1987). Butanols - four isomers: 1-butanol, 2-butanol, tert-butanol, isobutanol. World Health Organization. https://apps.who.int/iris/handle/10665/37266
- Ema, M., Hara, H., Matsumoto, M., Hirose, A., & Kamata, E. (2005).Evaluation of developmental toxicity of 1-butanol given to rats in drinking water throughout pregnancy. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association, 43(2), 325–331. https://doi.org/10.1016/j.fct.2004.11.003
- Köktürk, M., Alak, G., & Atamanalp, M. (2020).The effects of n-butanol on oxidative stress and apoptosis in zebra fish (Danio rerio) larvae. Comparative biochemistry and physiology. Toxicology & pharmacology : CBP, 227, 108636. https://doi.org/10.1016/j.cbpc.2019.108636